Die im Salz- oder Süßwasser lebenden Muscheln (Bivalvia) bilden innerhalb der Mollusken (Mollusca) oder Weichtiere eine Klasse und sind ein Beispiel für die Evolution im Tierreich. Sie kommen seit dem Kambrium in mit Schwankungen steigender Arten- und fossil dokumentierter Individuenzahl vor. Ihre Einteilung ergibt sich beispielsweise nach Bieler & Mikkelsen, 2006 (nur rezente Gruppen), Amler et al., 2000 (Einschluss auch von ausschließlich fossilen Gruppen), verschiedenen älteren Autoren (Thiele, 1935; Franc, 1960; Newell, 1965) sowie dem Zoological Record wie folgt (siehe auch Wikipedia):
Klasse Bivalvia Linnaeus, 1758
in Klammern Fotos (Seite.Nr.)
Die Reihenfolge der Fotos entspricht mit kleineren Umstellungen weitgehend der von Raymond C. Moore (Editor), Treatise on Invertebrate Paleontology, Part N, Mollusca 6, Bivalvia, (1969), The Geological Society of America and The University of Kansas. Die Veneroida wurden von mir hier an den Schluss gestellt, da sie mit einigen Ausnahmen von Familien anderer Ordnungen unter den rezenten Muscheln dominieren.
Die Fotos zeigen einige ausgewählte häufigere fossile und rezente Arten. Von Interesse sind aber auch die selteneren Formen, da deren Vorläufer teilweise in der relativen Vergangenheit wesentlich häufiger auftraten, was zur Frage ihrer Reduzierung führt, oder weil aus ihnen in der relativen Zukunft die dominierenden Spezies wurden beziehungsweise möglicherweise entstehen werden.
Biostratonomie
Myophoria kefersteini v. MÜNSTER aus dem Karnium Europas
Die Gattung Myophoria hat in der Trias, insbesondere im Germanischen Triasbecken bis in die Tethysbereiche, deren Sedimente heute in den Südalpen gefunden werden, eine weite Verbreitung. Zu den interessantesten Formen gehört Myophoria kefersteini v. MÜNSTER, die in beiden genannten Bereichen im unteren Keuper beziehungsweise Karn(ium) zu finden ist. Sie unterscheidet sich von der älteren und deutlich kleineren Myophoria vulgaris (v. SCHLOTHEIM) aus dem Muschelkalk vor allem durch ihre größeren Maße. M. SCHMIDT zeigt in Fig. 449 Myophoria kefersteini v. MÜNSTER aus der Bleiglanzbank des Gipskeupers (unt. Karn) von Sindelfingen. Die unten beigefügten Abbildungen stellen unterschiedliche Formen aus der Raibl-Gruppe der Südalpen dar. Diese Schichten bedecken in den Dolomiten den Schlern-Dolomit beziehungsweise die Cassianer und Wengener Schichten und werden ihrerseits vom Dachstein-Dolomit oder Hauptdolomit überlagert. Sie zeigen sich zum Beispiel an der Sella-Gruppe (siehe Bild) als deutlich abgezeichnete Trennebene, auf welcher der Hauptdolomit liegt, und kommen auch in den Lombardischen Alpen vor. Ähnliche Raibler Schichten finden sich im Norden in den Bayrischen beziehungsweise Tiroler Alpen über dem Wettersteinkalk und mit dem Dachsteinkalk oder Hauptdolomit im Hangenden.

Sella/ Dolomiten von der Marmolada aus gesehen (1958)
Die Raibler Schichten, die nach dem italienischen Ort Raibl (italienisch Cave del Predil, slowenisch Rabelj) mit einem ehemaligen Bleibergwerk nahe der Slowenischen Grenze benannt wurden, zeichnen sich durch ihren Fossilreichtum aus, der auf eine Bildung in nicht allzu großen Tiefen hinweist. Mächtige Riffe aus kalkabscheidenden Organismen entstehen bei langsamer Senkung des Meeresbodens. Dieser Prozess muss bei der Bildung der Raibler Schichten durch Absenkung oder eine erhöhte verfügbare Wassermenge beziehungsweise Verschlechterung der Lebensbedingungen für die Rifforganismen (Sedimente, Temperatur, Blei, ...) abrupt unterbrochen worden sein (Raibl Ereignis).
Das Auftreten von Myophoria kefersteini v. MÜNSTER oder ähnlichen Formen in den genannten Bereichen weist auf Meeresverbindungen zwischen den Vindelizischen Inseln hin. Im Süden fanden die Myophorien im Karn offensichtlich bessere Lebensbedingungen als im Keuper im Norden, wo sie im Muschelkalk, das heißt bis zum Ladin zahlreich vertreten waren.
E. RÜBENSTRUNK: Beitrag zur Kenntnis der deutschen Trias-Myophorien. Mitt. Großh. Bad. Geol. Landesanstalt, 6., S. 85-248, Heidelberg 1912, (Diss. 1909, C. Winter, Heidelberg).
M. SCHMIDT: Die Lebewelt unserer Trias. Rau, Öhringen 1928, 1939 Nachtrag I.
 
Myophoria kefersteini v. MÜNSTER, Raibl
 
Myophoria kefersteini v. MÜNSTER, Raibl Schafhaeutlia mellingi (HAUER), Raibl
 
Myophoria cf. kefersteini v. MÜNSTER, Gorenja vas, Slowenien
Stuoreswiesen bei St. Cassian, Dolomiten (1958)
An den Stuoreswiesen bei St. Cassian haben viele Paläontologen und Geologen bereits im 19. Jahrhundert in den Cassianer Schichten (Karn) mit Trachyceras aon (MÜNSTER) gesammelt.

Stuoreswiesen b. Corvara/ Dolomiten, Cassianer Schichten (1958)
Die Geologische Bundesanstalt in Wien verfügt über eine große Zahl von Belegstücken:
SIEBER, R.: Verzeichnis der Typus- und Abbildungsstücke der Geologischen Bundesanstalt;
5. Teil: Bivalvia der Trias (Zu Arbeiten von ALEXANDER BITTNER 1892-1902 und LUKAS WAAGEN 1907).-
Jb. Geol. B.-A., Band 126, Heft 3, S. 417-426, Wien 1984.
Die Bearbeitung der Bivalvia wurde insbesondere von A. Bittner und L. Waagen durchgeführt:
BITTNER, A.: Lamellibranchiaten der alpinen Trias. I. Teil: Revision der Lamellibranchiaten von Sct. Cassian. -
Abh. Geol. A.-A., 18, 1, S. 1-236, 24 Taf., Wien 1895.
WAAGEN,L.: Die Lamellibranchiaten der Pachycardientuffe der Seiser Alm nebst vergleichenden paläontologischen
und phylogenetischen Studien. (Fortsetzung, II. Teil, zu A. BITTNER, Lamellibranchiaten der alpinen Trias). -
Abh. Geol. A.-A., 18, 2, 1895-1907, S. 1-180, 19 Abb., 10 Taf., Wien 1907.
Über die Kleinwüchsigkeit der Fossilien wurde intensiv diskutiert.
Nachfolgend werden einige typische Muscheln der Cassianer Schichten gezeigt, die nur einen sehr kleinen Ausschnitt der umfangreichen Fauna darstellen.
Nucula strigilata GOLDF. - BITTNER 1895, p. 137, T. 17/
1-15,17 - Karn; S. Cassian - A (16) [vgl. DIENER
1925, T. 8/3a-c = T. 17/11,16, R. LEONARDI 1948, p.
41, T. 7/7-15. Palaeonucula st. (G.) - ZARDINI 1981, T.
1].

Palaeonucula strigilata (GOLDF.), Stuoreswiesen b. St. Cassian
Palaeoneilo lineata GOLDF. - BITTNER 1895, p. 133, T. 16/
1-11,14-16 - Karn; S. Cassian - A (13) [DIENER
1925, T. 8/2a-c = T. 16/13,16, R. LEONARDI1948, p.
44, T. 7/26-27]. Prosoleptus lineata (GOLDF.) - MOORE
N, p. 233, Fig. A 5/4a-b.

Prosoleptus lineata (GOLDF.), Stuoreswiesen b. St. Cassian
Cardita crenata GOLDF. - BITTNER 1895, p. 34, T. 41
5-10,11 (= LAUBE 1866, T. 15/12),12,13 - Karn;
S. Cassian - A (9) [KUTASSY 1931, p. 390. Palaeocardita crenata (MÜNST.) - MOORE
N, p. 554, Fig. E 54/1a-d.
 
Palaeocardita crenata (MÜNST.), Stuoreswiesen b. St. Cassian
Cassianella Beyrichii BITTNER 1895, p. 54, T. 6/16,18,20 -
Trias; S. Cassian - S (3)* [KUTASSY 1931, p. 270.
GUGENBERGER V/1935, p. 246. C. beyrichi B. - MOORE
N, p. 312, Fig. C 43/4a-c = BITTNER, T. 6116, R. ZARDINI
1981, T. 13].
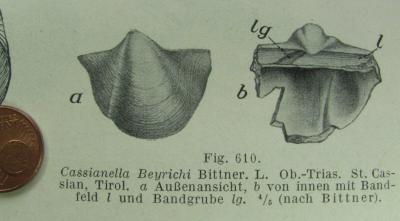
(aus Zittel I)
Formentera/ W-Mittelmeer (Juni/ Juli 1975)
Die folgenden Bilder zeigen Muschelfaunen der N-Küste zwischen Pujols und Es Caló sowie der S-Küste "Platja de Mitjorn" von Formentera. Auffällig sind die meist geringe Größe der vielfach juvenilen Individuen sowie die Unterschiede der beiden Küsten. Die Bilder erinnern allgemein an die Muschelfauna der Stuoreswiesen, wenn auch Unterschiede zu sehen sind, die schon durch den Evolutionsunterschied zwischen der Trias und der heutigen Zeit begründet sind.
 
Formentera N-Küste Cardita calyculata (L.)
 
Formentera S-Küste Cardita calyculata (L.) (untere Reihe)
Verschiedene Mittelmeerfaunen
 
Hammamet/ Tunesien 1976 Karthago, alter Kriegshafen, Ausgrabung UNESCO
 
Malia/ Kreta 1982 Psili Ammos/ Samos 2000
Rio Torsero bei Ceriale/ Riviera/ Mittelmeer (1959)
Die Schichten des Pliozän zeigen am Rio Torsero eine subtropische Muschelfauna (Länge der Sammlungsschachtel unten links 9 cm).
EUGENIO ANDRI, ANDREA TAGLIAMACCO, MASSIMILIANO TESTA, ANTONIO MARCHINI: Le Malacofaune fossili del Rio Torsero, Nuova Editrice Genovese 2005.
 
Rio Torsero, untere graue Lagen
Wangerooge/ Wattenmeer/ Nordsee (Mai 1996/7), Ostsee und Biscaya
Eine ganz andere Muschelfauna zeigt sich in der Nordsee an den Stränden der Insel Wangerooge. Bei den auf dem ersten Bild unten an der linken Seite abgebildeten Muscheln handelt es sich um Venerupis senescens (Cocconi) aus dem interglazialen Eem-Meer des Pleistozäns; die restlichen Muscheln sind rezent. Ein Vergleich dieser (realistischen) Fauna mit der von ZIEGELMEIER beschriebenen zeigt, dass dort viel mehr Arten genannt werden, die entweder an anderen Stellen oder nur sehr selten vorkommen. Bezogen auf geologische Verhältnisse legt dies nahe, dass Muscheln ein empfindlicher Umweltindikator sein können. Beispielsweise findet sich an den Stränden von Terschelling selten die Muschel Dosinia exoleta (L.), die ich im Wattenmeer noch nie gesehen habe und die beispielsweise in Arcachon häufig anzutreffen ist. In diesem Gebiet der Biscaya sind auch die Miesmuscheln viel größer als in der Nordsee oder gar in der Ostsee. Pectiniden finden sich beispielsweise in Arcachon massenhaft und an den Stränden der Niederlande sehr selten. So sind bereits in der Nordsee und der Biscaya, die durch den Kanal miteinander verbunden sind, in Abhängigkeit von den Umweltbedingungen große Unterschiede in der Muschelfauna zu beobachten.
B. ENTROP: Schelpen vinden en herkennen. 2. Auflage, N.V. W. J. Thieme & Cie, Zutphen, 1965.
E. ZIEGELMEIER: Die Muscheln (Bivalvia) der deutschen Meeresgebiete. Biologische Anstalt Helgoland, Nachdruck Hamburg 1962.
F. NORDSIECK: Die europäischen Meeresmuscheln (Bivalvia). Gustav Fischer Verlag, Stuttgart 1969.
 
Muscheln Wangerooge Dosinia exoleta (L.), Arcachon
 
Mytilus edulis L., Haffkrug/ Ostsee Mytilus edulis L., Brantevik/ Schonen/ Ostsee
 
Mytilus edulis L., Mimizan Plage/ Biscaya Chlamys varia (L.), Arcachon
Rotes Meer (um 1960)
Im Vergleich mit dem Mittelmeer kann die Fauna des Roten Meeres als tropisch bezeichnet werden. Das ist aus drei Gründen besonders überraschend. Zum einen beträgt der Abstand zum Mittelmeer weniger als 200 km, so dass die klimatischen Unterschiede hier eher gering sind; zum anderen wurde das Rote Meer im Tertiär gebildet und erhielt seine jetzige Form erst vor geologisch kurzer Zeit; außerdem ist es über 2000 km lang, und die Verbindung zum Indischen Ozean ist relativ schmal, so dass die Ausbreitung der Organismen erschwert war. Die Gründe für die faunistischen Unterschiede mögen in den Eiszeiten liegen, in denen der Mittelmeerraum vermutlich stärker abkühlte; weitere Gründe können im Einfluss der Zivilisationen gesucht werden, durch die das Rote Meer eher weniger beeinflusst wurde, wenn von der heutigen Schiffahrt und dem heutigen Tourismus einmal abgesehen wird.
Riesenmuscheln, wie die im ersten Bild unten gezeigte, treten zumindest bei Hurghada und Safaga heute häufig auf; ihre maximale Breite liegt nach eigener Beobachtung bei unter etwa 40 cm. Sie sind wie die übrige Fauna des Roten Meeres und insbesondere die Korallen streng geschützt.
P. G. OLIVER u. a.: Bivalved Seashells of the Red Sea. Verlag Christa Hemmen; National Museum of Wales, Cardiff, 1992.
 
Tridacna (Chametrachea) squamosa LAM. Modiolus auriculatus (KRAUSS)
 
Muscheln Hurghada
 
Muscheln Hurghada Muscheln Makadi Bay
Pholadomya (P.) transversa SEEBACH aus den Polyplocus-Schichten von NW- Deutschland
Die Art kommt insbesondere im Ton der Polyplocus-Schichten (Dogger beta) und möglicherweise auch im Dogger alpha beziehungsweise gamma von NW-Deutschland vor. Von Süddeutschland ist sie mir nicht bekannt, was mit unterschiedlichen Umweltbedingungen erklärt werden kann und die Trennung der beiden deutschen Jurameere durch eine Landmasse unterstreicht. Moesch erwähnt ein Exemplar von Salins/ Frankreich (?). Die Muschel erinnert an Pholadomya (P.) corrugata KOCH & DUNKER aus dem Lias sowie auch an die rezente Pholadomya (P.) candida G. B. SOW., eine der wenigen rezenten Formen der vom Jura bis zum Tertiär artenreichen Familie, deren Vertreter vermutlich im Schlick eingegraben lebten. Im Gegensatz dazu findet sich Inoceramus polyplocus ROEMER auch im südlichen deutschen Jurameer. Möglicherweise lebte diese Muschel, ähnlich wie Mytilus sp., auch angeheftet an Treibgut (siehe letztes Bild) und konnte so eine weitere Verbreitung finden.
Die abweichenden Formen, wie bei dem Exemplar von Dankersen, können vermutlich durch Verformungen im Ton erklärt werden und begründen wohl keine neuen Arten.
C. MOESCH: Monographie der Pholadomyen. Abh. der Schweiz. Paläont. Ges., Basel, 1874, 1875.
D. A. BRAUNS: Der mittlere Jura im nordwestlichen Deutschland ... . Verlag von Theodor Fischer, Cassel, 1869.
O. KUHN: Paläogeographie des deutschen Jura. Gustav Fischer Verlag, Jena, 1953.
 
Polyplocus-Schichten, S Wiehengebirge b. Bramsche
 
Polyplocus-Schichten, S Wiehengebirge b. Bramsche
 
Dogger beta/gamma?, Rinteln
 
Dogger beta/gamma?, Rinteln
 
Dogger beta/gamma?, Dankersen
 
Inoceramus (Mytiloceramus) polyplocus ROEMER, S Wiehengebirge b. Bramsche
Trigonia (T.) triangularis (GOLDF.) aus den Parkinsoni-Schichten von NW-Deutschland
Die Art ist typisch für das obere Bajocium in NW-Deutschland und kommt vor allem in einem breiten etwa westöstlichen Streifen von Osnabrück bis zum Hils vor. Sie wird von Lebküchner im südlichen deutschen Jurameer nur von wenigen Fundorten, wie zum Beispiel von Eningen beschrieben. Auch der Holotypus von Goldfuss stammt vermutlich von Lübbecke (zwischen Osnabrück und Minden). In Sengenthal fehlt die Art nach Weißmüller sowohl im Parkinsoni-Oolith als auch in den Garantianenschichten. Das unterschiedliche Vorkommen ist vermutlich eine Folge der verschiedenen Umweltbedingungen und unterstreicht die Trennung des deutschen Doggermeeres durch eine zentrale Landmasse. Die Art wird in Süddeutschland häufig verwechselt, zum Beispiel mit Trigonia interlaevigata QUENST..
Die Trigonien lebten insbesondere in der Jura- und Kreidezeit. Die einzige heute noch lebende Gattung der Familie ist Neotrigonia. Diese kommt insbesondere an den Küsten Australiens sowie Tasmaniens vor. Es wäre wohl voreilig, ohne nähere Prüfung aus den heute dort herrschenden Umweltbedingungen auf die Verhältnisse in vergangenen Meeren mit Vorkommen von Trigonien zu schließen.
LEBKÜCHNER, R.: Die Trigonien des süddeutschen Jura. Separat-Abdruck aus Palaeontographica Band LXXVII, E. Schweizerbart'sche Verlagsbuchhandlung, Stuttgart, 1932.
WEISSMÜLLER, M.: Die Muscheln des Bajocium von Sengenthal. Eigenverlag, 2009.
 
l. Kl. Gadderbaum b. Bielefeld r. Kl.
 
Gadderbaum b. Bielefeld
 
Velpe b. Osnabrück Bünde i. Westf.?
 
Velpe b. Osnabrück
 
Velpe b. Osnabrück
 
Velpe b. Osnabrück
 
Neotrigonia sp., Port Lincoln/S-Australien, rezent
Gotland/ Ostsee
Nach „Sveriges Geologi“ von N. H. MAGNUSSON ergibt sich im Südteil von Gotland von Nord nach Süd diese Schichtfolge aus dem mittleren und oberen Silur:
Hangendes
Hamra Kalkstein mit Pteronitella retroflexa (WAHLENBERG) u. a.
Burgsvik Schichten Pteronitella retroflexa (WAHLENBERG) u. a.
Eke Mergel
Hemse Gruppe mit Ilionia prisca (HISINGER) u. a.
Klinteberg Gruppe
Mulde Mergel mit Ptychopteria sowerbyi (McCOY) u.a.
Liegendes
Im „Lexique Stratigraphique International Vol. I, Europe, Fasciule 2c Suède“ von 1956 wird im Mulde Mergel u. a. Cornellites sowerbyi (McCOY) angegeben. Nach dem „Treatise on Invertebrate Paleontology part N Mollusca 6, Bivalvia“ von 1969 Seite N302 wird Cornellites WILLIAMS durch Ptychopteria HALL ersetzt. Diese Gattung sei stark prosoklin mit einem vorderen „Ohr“ und einem hinteren wohl entwickelten Flügel; die Zähne sollen stark variabel ausgebildet sein. Leptodesma (Seite N300 des „Treatise“) ist sehr ähnlich. Im Vergleich mit Pteronitella retroflexa (WAHLENBERG) (Seite N301 des „Treatise“) aus den Burgsvik Schichten und dem Hamra Kalkstein von Gotland oder der Öved-Ramsåsa Gruppe aus Schonen beziehungsweise von dem namengebenden Ludlow (Whitcliffe) ist das hier angeblich vorhandene actinodonte Schloss mit zahlreichen Zähnen kennzeichnend. Einen mehr oder weniger geschwungenen Ventralrand besitzen alle drei Genera; auch ist bei allen drei Gattungen die rechte Klappe etwas weniger konvex als die linke Klappe. Übrigens erinnert Hoernesia socialis (VON SCHLOTHEIM) aus dem (germanischen) Muschelkalk mit ihren nur angedeuteten Zähnen sowie den unterschiedlichen Klappen und der Schalenstruktur an Pteronitella retroflexa (WAHLENBERG).
Die von O. ISBERG in seinen „Studien über Lamellibranchiaten des Leptaenakalkes in Dalarna“ von 1934 beschriebene Muschelfauna stammt aus älteren Schichten (Ordovizium bis allenfalls unterstes Silur), so dass hier keine vergleichbaren Formen zu erwarten sind. Allerdings glaubt man in Ambonychinia oder anderen Gattungen die Vorfahren beziehungsweise Verwandte von Pteronitella zu erkennen.
Bereits JOACHIM BARRANDE diskutiert in dem Band „Acéphales“ seiner „Études locales et comparatives vol. VI“ von 1881 auf Seite 47 bis 48 das angebliche Schloss von Pteronitella, das er bei vergleichbaren Formen aus dem Silur und Devon in Böhmen nicht findet.
Die unten beigefügten Bilder zeigen fünf linke Klappen sowie eine mögliche rechte Klappe von Pteronitella retroflexa (WAHLENBERG) und eine Innenseite der linken Klappe von Ptychopteria sp. mit allenfalls leistenartigen Verdickungen im Schlossbereich sowie zur Vervollständigung ein Exemplar von Ilionia prisca (HISINGER) und zwei weitere Muscheln.
Das regionale Vorkommen von Pteronitella retroflexa (WAHLENBERG) kann biologische und/oder geologische Ursachen haben. Zum einen können die Lebensbedingungen im Bereich des heutigen Gotlands für die Art optimal gewesen sein; zum anderen kann hier auch ein relativ abgeschlossenes Becken ähnlich dem späteren (germanischen) Muschelkalkmeer mit schmalen Verbindungen nach Westen (Großbritannien, NE USA und Kanada) aber für diese Art schlechteren Verbindungen zur mitteleuropäischen Geosynkline vorgelegen haben.
V. G. WALMSLEY: The Identity and a New Description of Pteronitella retroflexa (WAHLENBERG) from the Upper Silurian of Gotland and the Welsh Borders. Geologiska Föreningen I Stockholm Förhandlingar, Vol. 84, 4, 1962, Seite 351-362.
E. PHILIPPI: Ueber das Schloss von Pterinaea retroflexa WAHLENB. sp.. Zeitschrift der Deutschen Geologischen Gesellschaft, Bd. 51, 1, 1899, Seite 181-183.
H. SHALER WILLIAMS & C. LEVENTHAL BREGER: The Fauna Of The Chapman Sandstone Of Maine; Including Descriptions Of Some Related Species From The Moose River Sandstone. Nabu Press 2012.
S. ELIASON: Fossilien auf Gotland. 2. Auflage, Gotlands Fornsal 2011.
 
Pteronitella retroflexa (WAHLENBERG), Kettelvik, Burgsvik Schichten, l. Kl.
 
Pteronitella retroflexa (WAHLENBERG), Kettelvik, Burgsvik Schichten, l. Kl.
 
Pteronitella retroflexa (WAHL.), Kettelvik l. Kl. Pteronitella retroflexa (WAHL.), N Burgsvik r. Kl.?
 
Ptychopteria sp., Djupvik, Mulde Mergel Ilionia prisca (HISINGER), Grogarnshuvud
Valar südl. Burgsvik, Burgsvik Schichten
Barrandium/ Prag
Insbesondere die Schichten des oberen Silur haben in der Umgebung von Prag eine reiche Muschelfauna geliefert, die 1881 von J. Barrande in seinem "Système silurien du centre de la Bohème", vol. 6 beschrieben wurde. Beispiele sind auf der Seite "Fotos 1.30 bis 1.43" zu finden. Hier soll nur die weit verbreitete Gattung Cardiola kurz dargestellt werden. Diese wurde in Europa von Schonen bis zu den Karawanken und von Tschechien in westlicher Richtung in Deutschland und Frankreich bis nach Nordamerika in Schichten des höheren Silur und Devon gefunden (in Nordamerika bis zum Unteren Karbon). Das Exemplar auf dem neunten Bild unten ist ein eiszeitliches Geschiebe und stammt vermutlich aus dem Colonus Schiefer von Schonen, der auch Pterochaenia glabra (GOLDF.) enthält. Nach dem bereits zitierten „Lexique Stratigraphique International Vol. I, Europe, Fasciule 2c Suède“ von 1956 enthält dieser unter anderem Cardiola interrupta (SOW.). Darüber liegt die Öved-Ramsasa Gruppe mit Pteronitella retroflexa (WAHLENBERG) in höheren Lagen und Palaeopecten danbyi (McCOY) in den zuunterst gelegenen Bjärsjölagard Schiefern, unter denen der Cardiola Schiefer der Colonus Schiefer anzutreffen ist. Danach sollten die Schichten mit Pteronitella retroflexa (WAHLENBERG) in Schweden jünger als die Schichten mit Cardiola interrupta (SOW.) im Barrandium sein, wobei natürlich nicht nur die Muscheln, sondern auch die übrige Fauna betrachtet werden müssen.
 
Cardiola interrupta (SOW.), Budnany Cardiola interrupta (SOW.), Kosor
 
Cardiola interrupta (SOW.), Lochkov
 
Cardiola interrupta (SOW.), Budnany Cardiola interrupta (SOW.), Jinonice
 
Cardiola bohemica BARR., Lochkov Cardiola bohemica BARR., Kosor

Cardiola cf. bohemica BARR., Großhansdorf b. Hamburg/ Schonen?, Colonus Schiefer (Geschiebe)

Palaeopecten ? sp., Bjärsjölagard/Schonen, Bjärsjölagard Schiefer
Daleiden/ Eifel (um 1980)
In den Wiltz Schichten des oberen Unterdevons treten Muscheln erstmals "massenhaft" mit großer Art- und Individuenzahl auf, wie zum Beispiel das erste Bild unten der kleineren Formen zeigt. Die Gründe dafür sind vermutlich optimale biologische Bedingungen und optimale Erhaltungsgegebenheiten. Die großen Monographien täuschen manchmal vor, dass zahlreiche Fossilien gefunden wurden, die aber lediglich das Ergebnis von jahrzehntelanger Sammeltätigkeit sind, wie schon O. ISBERG in seinen „Studien über Lamellibranchiaten des Leptaenakalkes in Dalarna“ schreibt.
Über devonische Muscheln liegen neben jüngeren Werken vor allem die alten Monographien von BEUSHAUSEN, FRECH und HALL vor. Da es sich bei deren Abbildungen um (mehr oder weniger subjektive) Zeichnungen handelt, ist wie bei BARRANDE eine gewisse Vorsicht angebracht. Andererseits können diese in manchen Fällen besser als Fotografien sein.
Die Auswahl der unten abgebildeten Muscheln stammt, soweit nicht anders bezeichnet, aus Daleiden.
L. BEUSHAUSEN: Die Lamellibranchiaten des rheinischen Devon mit Ausschluss der Aviculiden. Abhandl. der Königl. Preuss. geol. Landesanstalt, Neue Folge, Heft 17, 1895.
F. FRECH: Die devonischen Aviculiden Deutschlands - Ein Beitrag zur Systematik und Stammesgeschichte der Zweischaler. Abhandl. zur geologischen Specialkarte von Preussen und den Thüringischen Staaten, Band IX, Heft 3, 1891.
J. HALL: Lamellibranchiata I and II, descriptions and figures of the Monomyaria (I) and Dimyaria (II) of the upper Helderberg, Hamilton, (Portage,) and Chemung Groups. N. Y. Geol. Survey, Paleont., v. 5, 1884-1885.
CHR. FRANKE: Die Klerf-Schichten (Unter-Devon) im Großherzogtum Luxemburg, in der Westeifel (Deutschland) und im Gebiet von Burg Reuland (Belgien): fazielle und biostratigraphische Deutungen. Ferrantia 46, 2006, Seite 42-93.
 
Kleinfunde
 
Solemya ? compressa (GOLDF.)
 
 
Pterinea costata GOLDF. Ptychopteria (Actinopteria) lamellosa (GOLDF.)
 
Leptodesma (Leiopteria) sp.
 
Leptodesma (Leiopteria) arduennensis (STEINING.) Leptodesma (Leiopteria) arduennensis (STEINING.)
 
Gosseletia sp., Halenbach Gosseletia carinata (FOLLMANN)
 
Modiomorpha ? antiqua (GOLDF.) Paracyclas rugosa (GOLDF.)
Edmondia inflata (STEININGER), Kickeshausen Edmondia inflata (STEININGER)
 
Edmondia sp. Grammysia sp.
 
Grammysia anomala rhenana BEUSH. Grammysia cf. anomala rhenana BEUSH.
|


